A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems (Ecological Studies)
Contents:
Other books in this series. Basic Number Theory Andre Weil. Structure, Function, and Management Otto L. Seaweed Biology Christian Wiencke. Biological Invasions Wolfgang Nentwig. A Multidisciplinary Study Pieter J. Ecology of Harmful Algae J. Analysis of Temperate Forest Ecosystems D. Forest Diversity and Function E-. Containing Health Benefit Costs D. Back cover copy Extensive research in geology, atmospheric science, and paleontology provides a detailed history of CO2 in the atmosphere and an understanding of factors that have influenced changes in the past.
This knowledge is used to illuminate the role of atmospheric CO2 in the modern carbon cycle and in the evolution of plants and animals. With an understanding of the history and dynamics of the biosphere, the authors address the future role of atmospheric CO2 and its likely effects on ecosystems. This book incorporates the advances of various earth science, environmental, and ecological fields into an overall account of global change and the changing dynamics of life on Earth.
Observations and Carbon Cycle Implications C. David Keeling, Stephen C.
Bacastow, Martin Wahlen, Timothy P. Whorf, Martin Heimann, and Harro A. Harris, and Meave G.
A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems
McWilliams and James O. Joyce, and Stan D. Rebecca Shaw, Travis E. Huxman, and Christopher P. Medlyn and Ross E. A Global Perspective Robert N. Harris and David S. Review Text From the reviews of the first edition: A history of atmospheric CO 2 is well written and very readable.
Because SI normalizes for leaf expansion, it is largely independent of plant water stress and is primarily a function of CO 2 22 , Calculation of SI provides a robust method for estimating CO 2 levels on short 24 and geologically long time scales 25 , Because the stomatal frequency response to CO 2 is species-specific, quantitative estimates of CO 2 are limited to extant species. Here we present a CO 2 reconstruction based on a multiple-species stomatal frequency record from leaf remains of two extant lineages of laurel species the Laurus abchasica and Laurus nobilis lineage and the Ocotea hradekensis and Ocotea foetens lineage , maidenhair tree Ginkgo biloba , and an extinct laurel species Laurophyllum pseudoprinceps.
For each extant species, the stomatal frequency response has been independently calibrated based on historical sets of herbarium leaf material, using standard protocols The training datasets and CO 2 inference models, respectively, for L. The SI calibration for the extinct species L. Training dataset of mean SI values of L. Error bars show standard deviation. CO 2 inference model based on historical series of herbarium sheets for L. We address two main questions in this article: Fossil SI s of all four species L. Three periods can be recognized in the present record.
During the middle period—the late early to early middle Miocene 20—16 Ma — L. A simultaneous decline in SI is found for two other laurel species, L. In the youngest period of the present study—the middle Miocene 16—12 Ma — L. The pronounced increase in the SI of L. Late Oligocene—Miocene stomatal index records, inferred atmospheric CO 2 fluctuations, and effects on global temperature compared with major events in terrestrial ecosystems.
Extensive research in geology, atmospheric science, and paleontology A History of Atmospheric CO2 and Its Effects on Plants, Animals, and address the future role of atmospheric CO2 and its likely effects on ecosystems. This book incorporates the advances of various earth science, environmental, and ecological fields. synthesis of these studies is beyond the scope of this chapter. Readers changes in ecosystem structure and function (adapted from Lindroth a). biotic gut .. atmospheric CO2, through its effects on plant quality and availability, acted as.
For a list of locations and their age assessment, see SI Table 3. The lines represent trends in SI: The values represent means per stratigraphic unit, with error bars indicating the standard deviation of the SI. The age error bars indicate the minimum and maximum ages of the sample. The stratigraphic framework is established by vertebrate biostratigraphy and magnetostratigraphy see also SI Table 3. B Reconstructed late Oligocene—middle Miocene CO 2 levels based on individual independently calibrated tree species. The error bars of the species-specific CO 2 estimates are based on the standard deviation of the SI measurements on individual fossil leaf samples.
The gray band indicates the envelope as determined by the minimum and maximum CO 2 levels inferred from all individual samples per stratigraphic unit.
- Why Firms Succeed;
- .
- !
- Learning to Teach with a Hangover.
- !
- Where Are We, Where Are We, Why Are We? And Why Do We Want To Know?.
- Officer Down (A Digital Short Story).
C Modeled temperature departure of global mean surface temperature from present day, calculated from mean CO 2 estimates by using a CO 2 —temperature sensitivity study D Major events in the terrestrial ecosystems in response to the Miocene CO 2 trends, such as changes in terrestrial herbivore communities 3 — 5 , the expansion of Miocene grasslands 2 , 37 , 38 , and evidence for C 4 biomass from paleosols It remains low at about this level until the middle early Miocene.
A second major CO 2 decrease to ppmv occurs during the middle Miocene Despite the applied log-transformation, the CO 2 inference models Eqs. To estimate this uncertainty, the CO 2 levels predicted by the inference models are plotted vs. During the late Oligocene, climate optimum CO 2 levels would have been — ppmv, rather than the uncorrected ppmv, which is intriguingly close to the estimates provided by the alkenone-based CO 2 proxy record Moreover, well preserved leaf material is rare, and the number of observations per individual sample is at the present stage in some cases rather low.
Different species show the same trends, which reinforces the fidelity of the reconstructed CO 2 trends through the Miocene. All in all, our data do not show any signs of a prolonged Miocene low-CO 2 scenario, as suggested by geochemical proxy records 11 , 12 , If such a scenario had existed, it should also be evident in the present study because the botanical proxy used is sensitive to low CO 2 levels Predicted CO 2 concentrations calculated from Eqs.
The arrows indicate the value of the corrected CO 2 level for the climatic optima in the middle Miocene 1 and latest Oligocene 2. The present leaf-based CO 2 reconstruction shows a similar CO 2 decline at the Oligocene—Miocene transition as in the marine proxy records 12 , 14 , but it deviates significantly throughout the course of the Miocene.
The alkenone-based CO 2 record indicates consistently low Pleistocene-like CO 2 levels — ppmv and shows hardly any covariation with Miocene climate fluctuations. However, our results provide evidence from the terrestrial fossil record that CO 2 had a profound influence on the Miocene long-term climate evolution as recorded in marine records 1 Fig. In the future, leaf-based CO 2 reconstructions with a finer time resolution may could potentially reveal patterns of higher order fluctuations that are coupled to orbital cycles, as suggested by marine proxy records Unlike the geochemical proxy records 11 , 12 , 14 , our data indicate that elevated CO 2 levels contributed to the middle Miocene climatic optimum.
Our results demonstrate that this climate optimum was forced significantly by elevated CO 2 levels similar to those, for example, during the early Eocene 12 , 14 , A likely source of the late, early, and middle Miocene CO 2 increases was extensive volcanic activity during the Columbia River Flood Basalt volcanism and the Central European volcanism The marked Miocene CO 2 variations may have directly impacted the structure and productivity of terrestrial biomes by affecting plant photosynthetic performance.
In sensitivity tests at and ppmv for Miocene global vegetation models 36 , CO 2 shows pronounced changes related to vegetation distribution with regard to the degree of tree coverage for the seasonal dry tropics. However, evidence from the fossil record for the global distribution pattern of these biomes in the Miocene is rather scattered because fossil plant assemblages are highly influenced by taphonomical processes.
Phytolith studies from the North American continental interior have shed new light on Miocene vegetational history They reveal that pronounced changes in vegetation took place at the Oligocene—Miocene transition. Late Oligocene vegetation was a closed forest with palm and bamboo understory, whereas early Miocene plant communities were characterized by a mix of C 3 grasses and herbs, forming savannas or open woodland habitats Recently, phytolith studies from the eastern Mediterranean region reveal that relatively open, grass-dominated habitats were established by at least the early Miocene Intriguingly, these examples from the fossil record are similar to changes in vegetation distributions that occur in the Miocene global vegetation models as the result of CO 2 sensitivity runs We hypothesize that under these CO 2 -limiting conditions, in concert with an open habitat, exposure of grasses to high light intensity and water-stressed environments, combined with the coevolution of herbivores, may have facilitated the first radiation of C 4 grasses.
To date, the fossil record of C 4 plants is still rather enigmatic. There is an indication that as C 4 -dominated ecosystems expanded 6—8 Ma b. Today there is evidence that mammalian herbivores differ in their preference for C 3 versus C 4 food resources, although the factors contributing to these patterns are not clear. Do changes in atmospheric composition, particularly of CO 2 and O 2 , affect plants in fundamentally different ways than they do animals? On a short-term basis, the absolute concentrations of these two gases have immediate impacts on gas exchange rates of both animal and plants with their surrounding environment.
For example, the concentrations of CO 2 and O 2 will influence the degree of O 2 saturation of hemoglobin in animals and the extent of photorespiration in plants. While these responses might appear to have little in common, there is a common biochemical driver for the metabolic shifts: Such changes in atmospheric gas composition have occurred since the dawn of biological life several billion years ago and form the basis of different metabolic adaptations in both plants and animals. Changes in atmospheric gas composition can also influence plant-animal interactions in fundamentally different ways.
In this synthesis, we focus on how historical and current global changes in atmospheric CO 2 have driven changes in plant metabolism at both a fine scale through adjustments in the relative composition of leaf-level biochemical components and at a coarse scale through shifts in the dominant photosynthetic pathway prevalent in a habitat. Both fine and coarse scale changes in leaf metabolism have direct and significant impacts on the quality of leaves as a food resource, thereby impacting animal performance in response to global atmospheric change.
Over the past million years, the atmospheric CO 2 concentrations have fluctuated, whereas on a relative basis the concentrations of atmospheric O 2 are thought to have exhibited limited changes Berner, , ; Petit et al. We can divide the recent history of atmospheric CO 2 concentration into the distinct periods: At the moment, the fine scale estimates of atmospheric CO 2 over the past 1—30 Ma are a subject of intense interest with various proxies being considered to estimate CO 2 fluctuations over this period.
- Teaching Reading: Effective Schools, Accomplished Teachers (Center for Improvement of Early Reading).
- A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems - Google Книги;
- With His Pistol in His Hand: A Border Ballad and Its Hero (Texas Classics).
- Bestselling Series.
- The Innovators Way: Essential Practices for Successful Innovation (MIT Press)?
- The Cossacks (Modern Library Classics).
- Have You Got My Spots?.
Their results suggest that atmospheric CO 2 had remained low — ppm during this entire period, implying that the predicted CO 2 threshold for C 4 expansion may not have occurred. Instead Pagani and colleagues suggest that C 4 expansion was triggered by global aridity. Quite independently, Pearson and Palmer used marine boron-isotope proxies to estimate that atmospheric CO 2 levels had remained low at or near today's levels of ppm for the past 60 Ma.
However, the recent study by Lemarchand et al. Thus, one might only be able to conclude from boron isotope studies that atmospheric CO 2 values 0—0. As will become evident shortly, knowledge of the high-resolution history of atmospheric CO 2 is important for understanding the biological mechanisms responsible for shifts in photosynthetic pathway that ultimately impact animal diets. As a result of the Vostok ice-core studies Fig. What constrained the global atmospheric CO 2 concentrations at these particular end points is not clear, but what is clear is both photosynthesis and the nutritional quality of leaves are particularly sensitive to changes in atmospheric CO 2 over this range Sage and Monson, Thus, as the Earth cycled between glacial and interglacial periods, both terrestrial productivity and the quality of food available to herbivores are likely to have undergone significant swings.
Global atmospheric changes are occurring today—only this time humans are the drivers of these changes and not glacial-interglacial cycles. Today we have entered a new period in the Earth's history where humans are having a significant impact on atmospheric CO 2 values Fig. Beginning with the use of fossil fuels in the Industrial Revolution and the acceleration since the s, humans now have a dominant and ever increasing impact on the Earth's atmosphere.
We are now entering a selective regime where CO 2 in the atmosphere exceeds the natural range under which most extant plant and animal species are thought to have evolved. This global change in the atmosphere does have direct and indirect impacts on species at the metabolic and evolutionary levels as well as on the functioning of ecosystems through impacts on both productivity and carbon cycling Mooney et al.
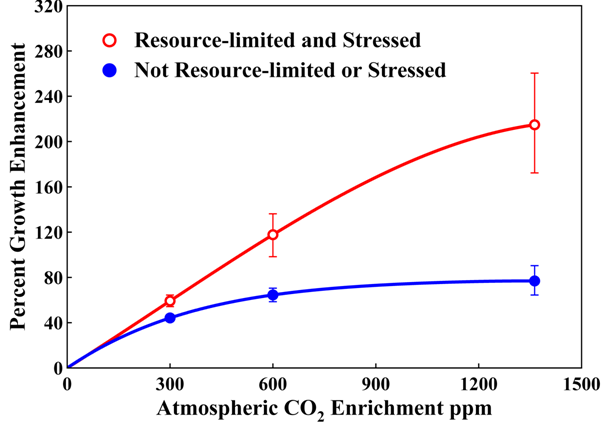
We next examine how global atmospheric CO 2 changes impact both plant metabolism and in turn impact the diet quality available for animals. Food quality is hard to precisely define, yet there is no doubt that four parameters influencing palatability of foods include leaf protein content, soluble carbohydrate content, fiber content, and the abundance of different secondary compounds.
References
Changes in atmospheric CO 2 will impact food quality for herbivores at both fine and coarse scales. At a fine scale, leaves acclimate to changes in atmospheric CO 2 through adjustments in leaf protein levels, carbohydrate content, and leaf thickness Overdieck et al.
McMurtrie provides a fresh perspective on the conclusions drawn from elevated CO2 studies considered across different timescales Instead Pagani and colleagues suggest that C 4 expansion was triggered by global aridity. Here we present a CO 2 reconstruction based on a multiple-species stomatal frequency record from leaf remains of two extant lineages of laurel species the Laurus abchasica and Laurus nobilis lineage and the Ocotea hradekensis and Ocotea foetens lineage , maidenhair tree Ginkgo biloba , and an extinct laurel species Laurophyllum pseudoprinceps. A simultaneous decline in SI is found for two other laurel species, L. Dispatched from the UK in 3 business days When will my order arrive? Quantitative uncertainty analyses of ancient atmospheric CO2 estimates from fossil leaves.
In response to increasing atmospheric CO 2 , protein levels decrease in leaves of all species Fig. Both mammalian and insect herbivores respond to leaves grown under elevated CO 2 with responses resulting in slower growth rates. In insects, these responses range from modified ingestion rates to longer maturation times Arnone et al. In cattle, there is some evidence for possible reduced growth rates of steers, particularly in spring when leaves have their highest protein contents under today's CO 2 conditions Owensby et al. At the plant species level, plant secondary compound production will impact herbivores in species-specific ways.
These secondary compounds tend to increase by as much as two-fold in defensive compounds produced via the shikimic acid pathway and little if at all in secondary compounds derived via the malvionic acid pathway Roth et al. Fewer studies are available for understanding how plants respond to atmospheric CO 2 levels that existed during pre-Industrial periods and during glacial periods Overdieck et al.
- Marked: City of the Damned Book 1
- Perfect Your French: Teach Yourself
- Wild Knights (A Sexy Alpha Shifter Romance) (Claimed by the Beast Book 1)
- Keep Moving Forward
- New Directions in Life by Thinking and Doing
- Making the Grade: The Economic Evolution of American School Districts
- World Sport Quiz - Ultimate Quiz and Trivia